Quelle est la différence entre l'alternative classique et la voie de la lectine
Le différence clé Entre alternative classique et voie de lectine est que l'initiation de la voie classique se déroule par la liaison des complexes antigènes-anticorps à la protéine C1Q, et l'initiation de la voie alternative se déroule par la liaison de C3B sur les surfaces étrangères, tandis que l'initiation de la voie de la lectine se déroule par la liaison au mannose lectine.
La voie du complément ou la cascade de complément fait partie du système immunitaire qui améliore la capacité des cellules phagocytaires et des anticorps à détruire et à effacer les microbes et à endommager les cellules d'un organisme, à favoriser l'inflammation et à attaquer la membrane cellulaire du pathogène. Les voies du complément sont générées par les systèmes immunitaires innés et adaptatifs. Ce système se compose de petites protéines qui sont synthétisées par le foie et circulent dans le sang sous la forme inactive. Ces protéines ou précurseurs sont activés dans les voies du complément. Il existe trois types de voies de complément: la voie classique, la voie alternative et la voie de la lectine.
CONTENU
1. Aperçu et différence clé
2. Qu'est-ce qu'une voie classique
3. Qu'est-ce qu'une voie alternative
4. Qu'est-ce qu'une voie de lectine
5. Similitudes - alternative classique et voie de lectine
6. Classical vs alternative vs voie de lectine sous forme tabulaire
7. Résumé - Classical vs alternative vs lectine Pathway
Qu'est-ce qu'une voie classique?
La voie classique est l'une des trois voies qui activent le système de complément. Le système de complément fait partie du système immunitaire. Les complexes antigènes-anticorps ainsi que les isotypes d'anticorps IgG et IgM lancent le système de complément. Les cellules apoptotiques, les cellules nécrotiques et les protéines en phase aiguë activent également la voie classique.
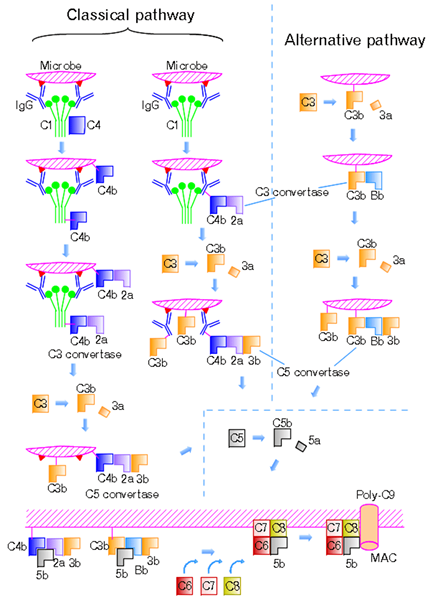
Figure 01: voie classique et voie alternative
Cette voie est initiée par la liaison des complexes antigènes-anticorps avec la protéine C1Q; La région globulaire de C1Q reconnaît et se lie à la région FC des anticorps IgG et IgM isotypes. Ils se lient également aux protéines de surface bactériennes et virales, des cellules apoptotiques et des protéines en phase aiguë. Pendant l'absence de facteurs d'activation, C1Q devient une partie du complexe C1 inactif, qui se compose de six molécules de C1Q, de deux molécules de C1R et de deux molécules de C1S. La liaison de C1Q conduit à des changements conformationnels et à l'activation de la sérine protéase C1R. Cela active et clive la sérine protéase C1S. C1S Clive C4 en C4A et C4B et C2 en C2A et C2B. C4B aide à la formation de C3 Convertase, C4BC2A. La C3 Convertase a la capacité de cliver C3 en C3A et C3B, ce qui est un facteur important pour la prochaine réaction enzymatique. C3V se lie à C3 convertase en formant C5 convertase, C4B2A3B, tandis que C3A recrute des cellules inflammatoires à travers. Ceux-ci sont appelés anaphylatoxines. C5 convertase clive C5 en C5 A et C5B. C5B se combine avec d'autres composants terminaux pour former le complexe d'attaque membranaire (MAC). Cela conduit à la lyse des bactéries invasives par insertion dans les membranes des cellules cibles, créant des pores fonctionnels.
Qu'est-ce qu'une voie alternative?
La voie alternative est l'une des trois voies qui opsonisent et détruisent les agents pathogènes. Le virus, les champignons, les bactéries, les parasites, l'immunoglobuline A et les polysaccharides activent la voie alternative et forment un mécanisme de défense essentiel indépendant du système immunitaire. La protéine C3B déclenche cette voie, et cette protéine se lie directement à un microbe. Les matériaux étrangers et les tissus endommagés déclenchent également la voie alternative. Étant donné que C3B est libre et abondant dans le plasma, il a la capacité de se lier à une cellule hôte ou à une surface de pathogène. Différentes protéines régulatrices participent à la prévention de l'activation du complément sur la cellule hôte.
Le récepteur du complément 1 (CR1) et le facteur d'accès à la désintégration (DAF) rivalisent avec le facteur B pour se lier à C3B à la surface de la cellule et éliminer BB du complexe C3BBB. Cliage de C3B dans la forme inactive, IC3B, par la protéase plasmatique appelée facteur de complément 1 inhibe la formation de C3 convertase. Le facteur de complément 1 nécessite un cofacteur de protéine de liaison C3B tel que le facteur H, CR1 ou le cofacteur membranaire de la protéolyse. Le facteur H inhibe la formation de C3 convertase en rivalisant avec le facteur B pour se lier à C3B. Cela accélère également la décroissance de la C3 Convertase. CFHR5, qui est la protéine 5 liée au facteur H complément.
Qu'est-ce qu'une voie de lectine?
La voie de lectine est un type de réaction en cascade dans le système de complément. Après l'activation de cette voie, l'action de C4 et C2 produit des protéines de complément activées plus loin dans la cascade. Cette voie ne reconnaît pas un anticorps lié à sa cible et initie avec la lectine de liaison au mannose (MBL) ou la liaison de la ficoline à certains sucres. Ce MBL se lie à des sucres tels que le mannose et le glucose avec des groupes OH en position terminale sur les glucides ou les glycoprotéines des bactéries, des champignons et certains virus.
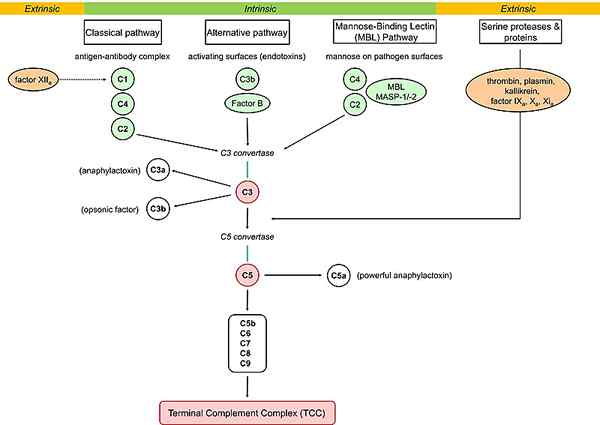
Figure 02: Pathways de complément
MBL, également connu sous le nom de protéine de liaison au mannose, est capable de lancer le système de complément en se liant aux surfaces des agents pathogènes. Les multimères de MBL forment des complexes avec des sérine protéases (la sérine protéase associée à la lectine de la lectine du mannose: MASP1, MASP2 et MASP3) qui sont des zymogènes protéiques. Ils sont similaires à C1R et C1 dans d'autres voies. MASP1 et MASP2 activent pour cliver les composants C4 et C2 en C4A, C4B, C2A et C2B. C4b a tendance à se lier aux membranes des cellules bactériennes. S'il ne s'active pas, il se combine avec C2A pour former C3 C3 Convertase opposant alternative C3 convertase. C4A et C2B agissent comme des cytokines puissantes. Le C4A provoque la dégranulation des mastocytes et des basophiles, et C2B augmente la perméabilité vasculaire.
Quelles sont les similitudes entre l'alternative classique et la voie de la lectine?
- Les voies classiques, alternatives et de lectine sont activées par une cascade de réactions menant au complexe d'attaque membranaire.
- Ils font partie du système immunitaire.
- Chaque voie a des protéines uniques pour l'initiation.
- Ils sont activés par certains isotypes d'anticorps liés aux antigènes
Quelle est la différence entre l'alternative classique et la voie de la lectine?
L'initiation de la voie classique a lieu à travers la liaison des complexes antigènes-anticorps à la protéine C1Q. L'initiation des voies alternatives se déroule à travers la liaison de C3B sur des surfaces étrangères, tandis que l'initiation de la voie de la lectine se déroule à travers la lectine de liaison au mannose. Ainsi, c'est la principale différence entre l'alternative classique et la voie de la lectine. Le rôle de la voie classique est qu'il agit comme le bras effecteur de l'immunité adaptative tandis que les voies alternatives et de lectine fonctionnent dans l'immunité innée. De plus, les activations C4 et C2 dans la voie classique sont C1S, et dans la voie de lectine est MASP-2, alors qu'il n'y a pas d'activation C4 et C2 dans la voie alternative.
L'infographie ci-dessous présente les différences entre l'alternative classique et la voie de la lectine sous forme tabulaire pour une comparaison côte à côte.
Résumé - Classical vs alternative vs lectine Pathway
L'initiation de la voie classique a lieu à travers la liaison des complexes antigènes-anticorps à la protéine C1Q. L'initiation des voies alternatives se déroule à travers la liaison de C3B sur des surfaces étrangères tandis que l'initiation de la voie de la lectine se déroule à travers la lectine de liaison au mannose. Donc, c'est la principale différence entre l'alternative classique et la voie de la lectine.
Référence:
1. «Path classique.«Biolabs créatifs.
2. Fujita, Teizo. «L'évolution de la voie de complément de lectine et de son rôle dans l'immunité innée.»Nature News, Nature Publishing Group.
Image gracieuseté:
1. «Pathways de complément."Par TOLSH_ENG - https: // téléchargement.wikimedia.org / wikipedia / communes / b / b0 / complément pathways.PNG (CC BY-SA 4.0) via Commons Wikimedia
2. "Figure 1.4. Activation du complément »par Hofman.Martijn - Propre travaux (CC BY-SA 4.0) via Commons Wikimedia